Kristen Krumhardt (Sep 18 2020 at 18:13): Kristen Krumhardt (Sep 18 2020 at 18:13):
Kristen Krumhardt (Sep 18 2020 at 18:13): Kristen Krumhardt (Sep 18 2020 at 18:13):Hi @Matt Long @Keith Lindsay @Michael Levy
I have been puzzling about issue with coccoCaCO3 that manifests only when I run with additional zooplankton groups.
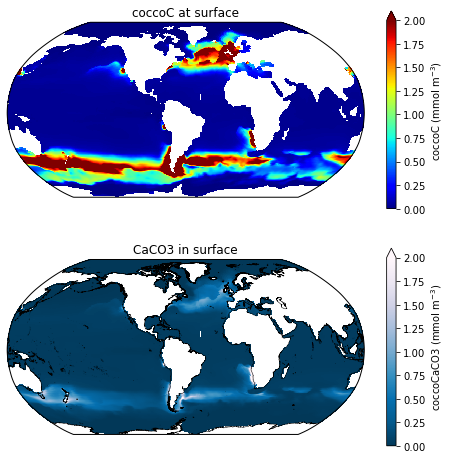
Here are the cocco biomass fields for a run with just one zooplankton (a normal cocco run): MARBL_4p1z_cocco.png
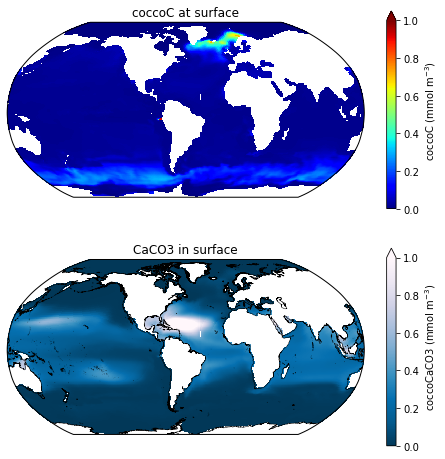
Here are the cocco biomass fields for a run with 4 zooplankton: MARBL_4p4z_cocco.png
There appears to be a disconnect between where most of the coccoC is and where the coccoCaCO3 is in the 4zooplankton run. These runs were done out of the same sandbox. I've chatted with Mike about this issue, but we can't seem to find any obvious reasons for the disconnect between coccoC and coccoCaCO3 in the runs with more zooplankton. The phytoplankton parameters in these runs are exactly the same, except for the QCaCO3_max is set to 1 in the 4zoo run and QCaCO3_max set to 2 in the 1zoo run (which is the default for an explicit calcifier). I had to make this change because, if I have QCaCO3_max=2.0 in the 4zoo run, it crashed with an error that said something about the ballast exceeding the POC flux...
Also worth noting, this pattern also shows up on the CaCO3_PROD_zint variable, but globally integrated calcification is still very low for the 4p4z run.. and surface Alk still has positive bias so the excessive coccoCaCO3 in mid to low latitudes doesn't seem to be projecting onto alkalinity.
My diagnostic notebooks on these two runs are on github:
The 4 zooplankton run:
https://github.com/kristenkrumhardt/MARBL_4p4z/blob/master/tuning_notebooks/MARBL_diagnostic_hist_4p4z.003.ipynb
The 1 zooplankton run:
https://github.com/kristenkrumhardt/MARBL_4p4z/blob/master/tuning_notebooks/MARBL_diagnostic_hist_cocco.001.ipynb
History files are here for the 4 zooplankton run:
/glade/scratch/kristenk/archive/g.e22a06.G1850ECOIAF.T62_g17.4p4z.003/ocn/hist
and for the 1 zooplankton run:
/glade/scratch/kristenk/archive/g.e22a06.G1850ECOIAF.T62_g17.4p4z.coccotest/ocn/hist/
The directory where I keep all these cases is here:
/glade/work/kristenk/cesm_work/cesm2.2_4P4Z_tuningcases/
If you have any ideas about what could be going wrong, please let me know.
Kristen Krumhardt (Sep 23 2020 at 20:10):Hi @Keith Lindsay @Matt Long @Michael Levy ,
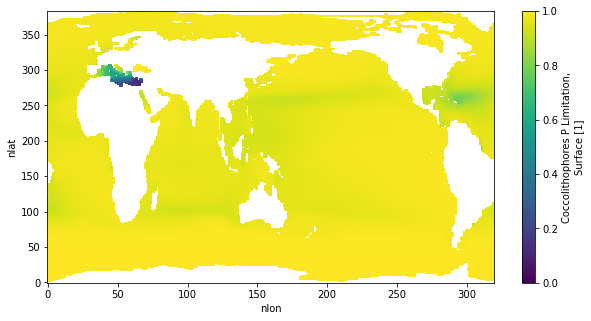
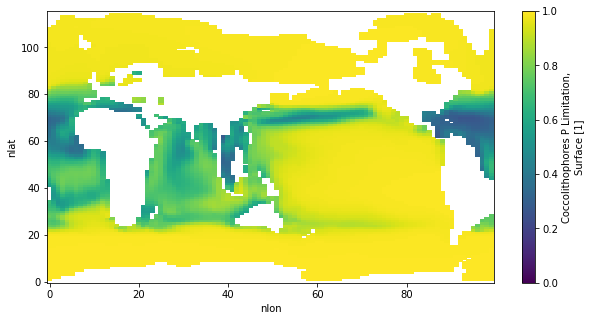
I ran a couple tests over the last couple days and think I figured out this coccoCaCO3 problem. The issue is that there is much more P-limitation in the 4 zooplankton set up. There is a well documented increase in coccolithophore PIC/POC ratios under P-limitation in lab studies. This was part of my coccolithophore parameterization, but it never really had much of an effect since there's not usually much P-limitation at all in CESM. However, when I added additional zooplankton, P limitation became much more severe. I think this is because there's more POC/POP sinking flux with more zooplankton. it caused my parameterization to have much more of an effect (perhaps unrealistic) than I've ever seen before.
Here is cocco P-limitation with only one zooplankton:
download-3.png
Here is cocco P-limitation with 4 zooplankton:
download-5.png
For now I can just comment out the P-limitation effect on coccolithophore PIC/POC.. then the coccoCaCO3 looks normal.
So for tuning the 4P4Z version, should I aim to reduce P-limitation? ... or is more P-limitation ok? (just not having such an extreme effect on coccolithophore PIC/POC)?
Precious Mongwe (Sep 29 2020 at 16:19):An iron cycle cascade governs the response of equatorial Pacific ecosystems to climate change
Tagliabue et al., 2020
Earth System Models project that global climate change will reduce ocean net pri-mary production (NPP), upper trophic level biota biomass and potential fisheries catches in the future, especially in the eastern equatorial Pacific. However, projec-tions from Earth System Models are undermined by poorly constrained assumptions regarding the biological cycling of iron, which is the main limiting resource for NPP over large parts of the ocean. In this study, we show that the climate change trends in NPP and the biomass of upper trophic levels are strongly affected by modifying assumptions associated with phytoplankton iron uptake. Using a suite of model ex-periments, we find 21st century climate change impacts on regional NPP range from −12.3% to +2.4% under a high emissions climate change scenario. This wide range arises from variations in the efficiency of iron retention in the upper ocean in the eastern equatorial Pacific across different scenarios of biological iron uptake, which affect the strength of regional iron limitation. Those scenarios where nitrogen limita-tion replaced iron limitation showed the largest projected NPP declines, while those where iron limitation was more resilient displayed little future change. All model sce-narios have similar skill in reproducing past inter-annual variations in regional ocean NPP, largely due to limited change in the historical period. Ultimately, projections of end of century upper trophic level biomass change are altered by 50%–80% across all plausible scenarios. Overall, we find that uncertainties in the biological iron cycle cas-cade through open ocean pelagic ecosystems, from plankton to fish, affecting their evolution under climate change. This highlights additional challenges to developing effective conservation and fisheries management policies under climate change.
https://onlinelibrary.wiley.com/doi/pdf/10.1111/gcb.15316
Precious Mongwe (Oct 10 2020 at 16:45):Optimality-based non-Redfield plankton–ecosystem model (OPEM v1.1) in UVic-ESCM 2.9 – Part 1: Implementation and model behaviour
Pahlow et al 2020
A implementation of an optimality-based plankton– ecosystem model (OPEM) version 1.1 with variable carbon : nitrogen : phosphorus (C : N : P) stoichiometry in the University of Victoria ESM (UVic; Eby et al., 2009; Weaver et al., 2001) and the behaviour of two calibrated reference configurations, which differ in the assumed temperature de- pendence of diazotrophs. Compared to the classic fixed-stoichiometry UVic model, OPEM is closer to recent satellite-based estimates of net community production (NCP), despite overestimating net primary production (NPP), can better reproduce deep-ocean gradients in the NO− : PO3− ratio and partially explains ob- 34 served patterns of particulate C : N : P in the surface ocean. Allowing diazotrophs to grow (but not necessarily fix N2) at similar temperatures as other phytoplankton results in a bet- ter representation of surface Chl and NPP in the Arctic and Antarctic oceans. Deficiencies of our calibrated OPEM configurations may serve as a magnifying glass for shortcomings in global biogeochemical models and hence guide future model devel opment.
https://gmd.copernicus.org/articles/13/4663/2020/gmd-13-4663-2020.pdf
Last updated: May 16 2025 at 17:14 UTC